
Modèle fruitweb Carpocapse
Téléchargez cette explication au format PDF
Le modèle est basé sur le modèle du carpocapse développé en Suisse par Graf, Höhn, Höpli et Kuske 2018. Il s’agit d’un modèle de population, c’est-à-dire que tous les stades commençant par les pupes du carpocapse au printemps sont simulés. Au début du modèle, on suppose 100 nymphes.
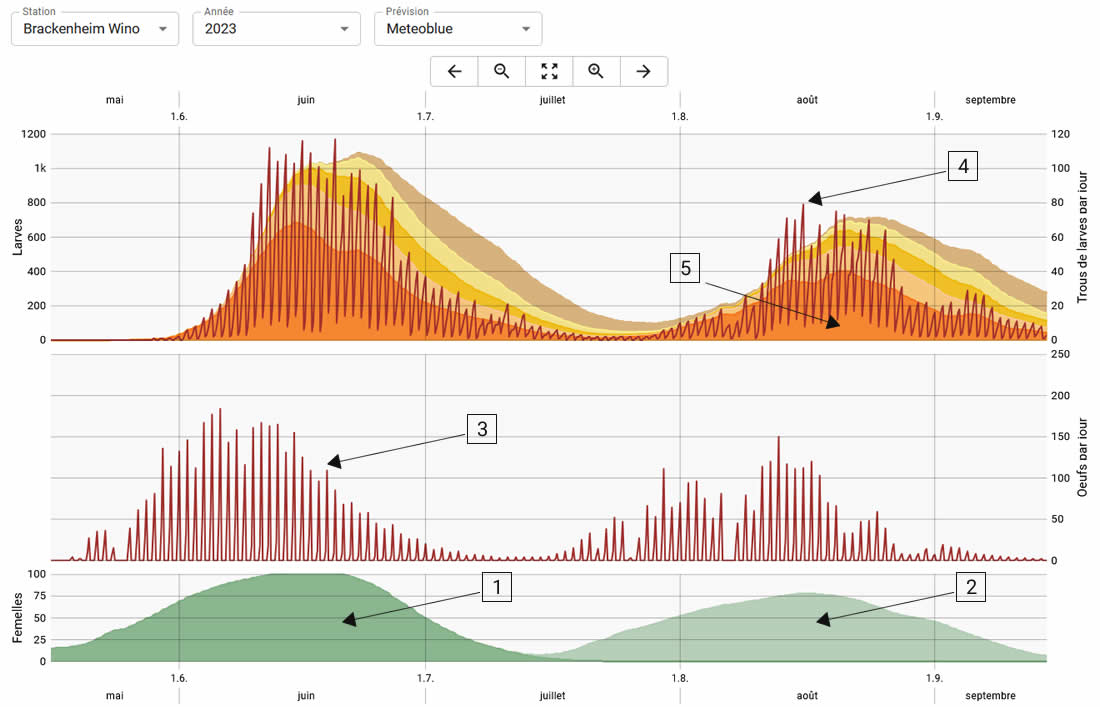
Une attention particulière a été accordée à la ponte, aux larves L1 et aux trous frais percés par les larves L1. Les vols, la ponte et les trous sont effectuées chaque jour en fonction des conditions climatiques. La présence des papillons (femelles) et les stades larvaires sont indiqués sous forme de somme et leur développement dans le temps.
Point de départ (Biofix) :
Si la date de première capture n’est pas encodée, le modèle commence à calculer les sommes de température à partir du 1er janvier (T0 > 10°C, valeur moyenne 417 DD, SD 63) pour l’éclosion des premiers carpocapses. Les pupes du carpocapse se développent à trois vitesses différentes. Cela dépend de l’exposition de la nymphe au soleil. Pour l’instant, le rayonnement solaire réel n’est pas encore utilisé pour calculer la vitesse de développement, mais des paramètres théoriques sont pris en compte . Toutefois, cette méthode sera bientôt remplacée par celle publiée par Graf et. al. 2003, qui inclut le rayonnement solaire.
Les premiers tests ont montré que le calcul du démarrage des vols avec cette méthode fonctionne très bien. Cela est particulièrement vrai pour la région du sud de l’Allemagne et de l’Alte Land. S’il devait y avoir des différences dans d’autres régions, il est possible de démarrer le modèle manuellement en saisissant le début du vol (piège à phéromones). Les premiers papillons capturés dans la région sont décisifs. Veuillez donc relever vos pièges ou demander à vos conseillers de vous communiquer cette date.
Le vol des femelles est représenté en bas du graphique. Les femelles de la première génération sont représentées par une zone vert foncé (1) et les femelles de la deuxième génération sont représentées par une zone vert clair (2). Actuellement, seules deux générations sont simulées.
Fécondité des femelles
Selon les études de Graf et al. 2018, la fertilité est maximale au début de leur activité de ponte (après la phase de pré-oviposition) et diminue ensuite de manière significative avec le temps. Graf et. al. 2018 travaillent avec une distribution de Weibull. Nous avons fait une approximation avec une distribution d’Erlang (To > 10°C, valeur moyenne 115 DD, SD 66).
Dépôt des oeufs
Les femelles du carpocapse pondent leurs œufs le soir, au crépuscule. L’intensité de la ponte dépend de 3 facteurs
- Moment de la journée : les œufs sont pondus avant et après le coucher du soleil. Pour l’intensité de l’activité de ponte dans le temps, nous avons utilisé une distribution normale. L’intensité la plus élevée se produit au moment du coucher du soleil (valeur moyenne 2 heures, SD 0,5 heures).
- Conditions météorologiques : l’intensité de la ponte dépend de la température. En dessous de 14°C, il n’y a pas de ponte. La température optimale est de 23 à 25°C. Au-delà de 31 °C, aucun œuf n’est pondu. En cas de pluie, la ponte est interrompue.
- Fécondité : voir ci-dessus
Le graphique montre toujours la ponte par jour (3). Il est ainsi très facile de voir dans quelles phases la ponte est particulièrement élevée ou faible.
Développement des œufs et des larves
Les paramètres suivants sont utilisés pour le développement des œufs et les différents stades larvaires
| Stade de développement | Valeur moyenne DD | SD en % | Seuil de développement To | Distribution |
|---|---|---|---|---|
| Oeufs | 78 | 10 | 10°C | Erlang |
| L1 Larves | 45 | 10 | 10°C | Erlang |
| L2 Larves | 45 | 10 | 10°C | Erlang |
| L3 Larves | 45 | 10 | 10°C | Erlang |
| L4 Larves | 55 | 10 | 10°C | Erlang |
| L5 Larves | 120 | 10 | 10°C | Erlang |
| Pupes | 159 | 6 | 10°C | Erlang |
Chaque étape a été soumise à deux vitesses de développement. D’une part, la somme des températures a été calculée sur la base de la température de l’air mesurée, et d’autre part, un supplément a été calculé pour la partie de la population qui est exposée à un rayonnement solaire plus élevé (analogue au développement post-diapause).
Éclosion des larves
L’éclosion des larves est indiquée sur la ligne supérieure sous forme d’éclosion larvaire par jour (4). De plus, le nombre de cinq stades larvaires peut être représenté par des zones de couleurs différentes. Le stade larvaire le plus important L1 est indiqué en orange en bas (5).
Méthode de calcul
Le modèle de carpocapse a été développé sur la base du modèle de « time varying distributed delay model » (Manetsch 1976). Cela permet de calculer le développement de chaque individu simulé par intervalle de temps. Par conséquent, les transitions entre les différents stades (œuf, larve, pupe et adulte) ne sont pas normalement distribuées mais suivent une distribution d’acquisition.
Litérature
T.J. Manetsch (1976). Time-varying distributed delays and their use in aggregative models of large systems. IEEE Transactions on Systems, Man, and Cybernetics 6: 547–553.
B. Graf, H. Höhn, H.U.Höpli und S. Kuske (2018). Predicting the phenology of codling moth, Cydia pomonella, for sustainable pest management in Swiss apple orchards. The Netherlands Entomological Society Entomologia Experimentalis et Applicata 166: 618-627
B. Graf, H.U. Höpli und H. Höhn (2003). Optimizing insect pest management in apple orchards with SOPRA. Bulletin IOBC/SROP, Vol.26 No.11:43-48